Présentation
Les pelouses à Gispet (Festuca eskia) sont localisées sur des zones fraîches souvent en versants nord et ouest, principalement à l’étage subalpin, où la couche de neige tend à persister ce qui limite l'impact des variations de températures (phénomènes de gel-dégel) responsables de l’érosion des sols. En conséquence, elles ne sont généralement pas disposées en gradins (présence récurrente de zones érodées de manière horizontale), phénomène caractéristique d’autres pelouses à Gispet mieux exposées, ne relevant pas de ce type d’habitat Ces pelouses dites « fermées », relativement denses et drues, d'un vert foncé et de faible diversité floristique abritent en moyenne 10 à 15 espèces et sont généralement dominées par le Gispet et ou le Nard raide (Nardus stricta).
Ces formations sont relativement stables car les fortes contraintes imposées par la durée d'enneigement et le poids de la neige limitent la dynamique de colonisation par les ligneux. On peut observer des mosaïques de pelouse-lande à plus basse en altitude mais la part n’est pas toujours facile à faire entre une mosaïque spatiale, liée à des conditions de milieux différentes, et une mosaïque temporelle liée à une avancée de la lande (à Rhododendron et Myrtille essentiellement) sur la pelouse.
En France, ce type de pelouse est endémique de la chaîne des Pyrénées. Les pelouses acides à Festuca eskia, également appelées pelouses à Gispet, sont présentes sur l’ensemble des sommets siliceux, des Pyrénées-Orientales jusqu’au Haut-Béarn.
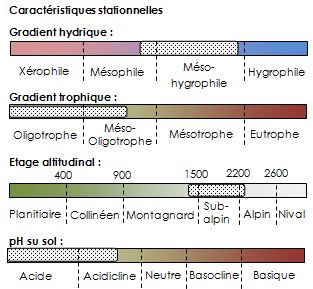
Ce groupement, que l’on trouve essentiellement à l’étage subalpin, a besoin d’une longue période d’enneigement qui le protège de l’action régulière du gel-dégel. Le substrat est composé d’un sol acide assez profond, caillouteux et humique, pauvre en éléments nutritifs. L’alimentation en eau est bonne, du fait de la période de fonte de la neige étalée dans la saison.
Cet habitat a une valeur patrimoniale importante compte tenu de son endémisme. Il n’est pas directement menacé sauf peut-être par le réchauffement climatique et à long terme par la déprise pastorale qui peut se traduire, dans certaines conditions, par une progression de la rhodoraie.
Gestion et conservation
 Exclusivement pâturées, les gispetières sont fréquentées par les troupeaux depuis le départ en végétation (juin-juillet) jusqu‘aux premières gelées (septembre-octobre). Compte-tenu de l’altitude et de la pente, elles sont plus représentées sur des quartiers ovins mais peuvent aussi être utilisées par les autres types de bétail.
Exclusivement pâturées, les gispetières sont fréquentées par les troupeaux depuis le départ en végétation (juin-juillet) jusqu‘aux premières gelées (septembre-octobre). Compte-tenu de l’altitude et de la pente, elles sont plus représentées sur des quartiers ovins mais peuvent aussi être utilisées par les autres types de bétail.
Malgré une importante biomasse, la valeur pastorale est faible à moyenne : 5 à 15 pour les gispetières denses [= indice de valeur pastorale calculé selon la méthode Daget & Poissonnet, 1971], 10 à 20 pour les faciès de transition avec les nardaies, plus diversifiées. Le Gispet est une graminée relativement tardive (floraison fin juillet à début août) mais ses limbes à croissance rapide deviennent rapidement durs et piquants. Tout comme le Nard (Nardus stricta), il est refusé ou à peine épointé par les brebis en tout début de saison, un peu mieux consommé par les bovins ou les équins. L’intérêt pastoral de ces pelouses tient surtout aux espèces compagnes, en particulier au Trèfle alpin (Trifolium alpinum), toujours présent dans les gispetières, bien qu’en quantités très variables. Diverses espèces fourragères - Agrostis capillaris, Meum athamanticum, Plantago alpina, Lotus corniculatus, Thymus serpyllum gpe… - contribuent également à la qualité pastorale de ces pelouses. Selon la date et les conditions d’exploitation (gardiennage, type de bétail, niveau de chargement), les animaux trient plus ou moins mais l’ouverture ainsi créée va toujours dans le sens d’une plus grande diversité floristique et d’une meilleure appétence.
En conditions de sous-pâturage, l’accumulation de litière non dégradée au niveau du sol accentue les difficultés d’exploitation de la pelouse. Dans la partie centrale et orientale des Pyrénées, il est fait mention d’anciens écobuages pour éliminer l’herbe sèche et « rajeunir » la pelouse. Cette pratique n’est plus en vigueur aujourd’hui.
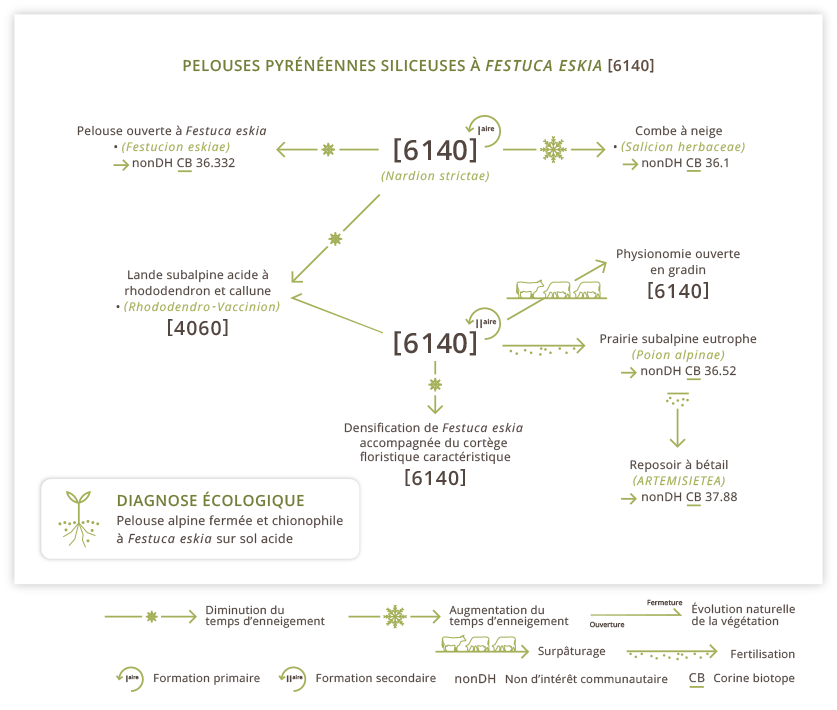
Déprise, abandon : Dans le contexte actuel du pastoralisme pyrénéen le scénario le plus probable ou le plus fréquemment évoqué est celui d’une diminution globale de la pression de pâturage sur les pelouses à Gispet. En cas d’abandon ou de sous-utilisation, on observe, en quelques années, une densification de la pelouse avec augmentation de la biomasse sur pied, et accentuation du phénomène de dominance par Festuca eskia.  La pelouse perd de sa valeur en devenant de moins en moins appétente et il est de plus en plus difficile d’y faire rester les animaux. Cette première phase d’évolution peut passer inaperçue car il s’agit de modifications internes de la flore [= la liste d’espèce reste pratiquement inchangée mais les contributions spécifiques et la physionomie évoluent] sans que l’on change de type d’habitat. Elle est cependant souvent déterminante pour son maintien à long terme.
La pelouse perd de sa valeur en devenant de moins en moins appétente et il est de plus en plus difficile d’y faire rester les animaux. Cette première phase d’évolution peut passer inaperçue car il s’agit de modifications internes de la flore [= la liste d’espèce reste pratiquement inchangée mais les contributions spécifiques et la physionomie évoluent] sans que l’on change de type d’habitat. Elle est cependant souvent déterminante pour son maintien à long terme.
A moyen et à long terme, cette situation de sous-pâturage chronique est propice au développement de ligneux bas : Rhododendron, Myrtille et Airelle ou Callune et Genévrier dans les conditions plus sèches. L’évolution vers la lande concerne principalement les altitudes intermédiaires (< 2200 m) avec une vitesse de fermeture qui reste à préciser : plus rapide dans les pentes fraîches inférieures à 2000 m où les landes à Rhododendron sont elles mêmes rapidement piquetées de sapins, de pins ou de bouleaux, elle reste difficile à appréhender quand les conditions de milieu sont moins favorables à la forêt.
Il est donc recommandé de maintenir une pression de pâturage régulière et précoce (avant épiaison) afin de limiter la densification du Gispet et de retarder le développement des ligneux. Si les conditions de pente et d’accès le permettent, les vaches et les chevaux sont moins sélectifs et un peu plus tolérants vis-à-vis de la phénologie.
Pression pastorale inadaptée : selon certains auteurs, les passages répétés de troupeaux ovins sur des gispetières denses de pente entraineraient l’ouverture de la pelouse et la création de gradins caractéristiques des gispetières ouvertes (hors Directive). Cette interprétation reste à confirmer.
Ponctuellement, les concentrations d’animaux (parcs de nuit, pierres à sel…) peuvent entrainer une transformation de la flore avec disparition, à terme, des espèces caractéristiques de l’habitat mais ce risque ne concerne que des surfaces très réduites.
Evolution climatique : La dominance de Festuca eskia et la physionomie caractéristique de ces pelouses denses dépendent en grande partie d’une couverture neigeuse prolongée qui limite les variations de température et favorise un bon degré d’humidité au printemps. Une réduction durable et répétée du niveau d’enneigement représenterait certainement une menace pour l’intégrité des pelouses les plus basses ou les plus exposées au vent et au soleil.
Schéma fonctionnel synthétique
Un schéma fonctionnel est une représentation théorique des trajectoires possibles de l'habitat en l'absence de gestion, en cas de changement de pratiques ou de conditions stationnelles.
La situation d'équilibre est obtenue par les modes de gestion proposés dans le paragraphe "gestion et conservation".
La diagnose écologique est un intitulé de l'habitat qui reprend les éléments stationnels et écologiques déterminants pour la définition de l'habitat (moins ambigüs que les intitulés des typologies codées).
Références et expériences de gestion
| massif pyrénéen | depuis 2013 | Mise en place d'un réseau de stations permanentes d'observation et de suivi pour évaluer et suivre l'état de conservation de l'habitat |
Mise en place d’un réseau permanent de placettes de référence : objectif final = 40 à 60 placettes couvrant la variabilité écologique, typologique et dynamique de l’habitat. Objectifs :
L'échantillon de stations est en cours de construction. |
contact : Catherine Brau-Nogué (CBNPMP) catherine.brau-nogue@cbnpmp.fr
|
| massif pyrénéen | 2013 | Réalisation d'un stage (M1) sur l'évaluation de l'état de conservation des pelouses pyrénéennes siliceuses à Festuca eskia. [Encadrement MNHN-CBN Méditerranéen] |
|
contact : Lise Maciejewski (MNHN) |
| Vallée de Gèdre-Gavarnie (65) | 2005 | Suivi de la progression et de la densification de pelouses à Festuca eskia | Mise en place d'un transect permanent en limite basse d'estive. Enregistrement tous les 2 ans de la présence et de la densité de Festuca eskia le long de ce transect. | contact : Parc National des Pyrénées |
BAUDIERE A., GESLOT A., CHIGLIONE C., NEGRE R., 1973. "La pelouse à Festuca eskia en Pyrénées centrales et orientales : esquisse taxinomique et écologique." Acta Bot. Acad. Sci. Hung., 19 (1-4) : 23-35.
Résumé auteur : Festuca eskia, a subalpine species of the pyrenean subalpine level, in reality actually consists in different types. One of them is characteristic of the geomorphogenic periglacial hill-sides, the other is characteristic of the biostatic habitats. The same is valid as regards the two types of Trifolium alpinum. Among identical conditions Campanula ficaroides opposes C. recta. As regards, Festuca paniculata, it is related to another type of Trifolium alpinum on the sub-alpine, high, cryoclastic hill-sides, while on the biostatic habitats it is related to Campanula precatoria. Therefore, the dynamics of the hill-sides remarkably indicates the speciation of the data, which are different both as regards their morphology, and their chemistry, that is their total amino-acid contents and proline in particular.
CLAUSTRES G., 1951, « Le rôle du Gispet (Festuca eskia) dans les pâturages ariégeois », Bulletin de la Société d'Histoire Naturelle de Toulouse, 86 : 31-35
Résumé CBN : l'article décrit la physionomie, la répartition et l'écologie des pelouses à gispet, plus particulièrement les pelouses ouvertes des fortes pentes de soulane. Il précise l'intérêt pastoral de ces formations pour les Pyrénées ariégeoises. Il mentionne d'anciennes pratiques de fauche (19e siècle) ainsi que quelques usages plus récents (litière, toiture, paillasse) du foin de gispet. L'article est aussi intéressant pour les informations toponymiques et étymologiques.
CLAUSTRES G., 1966, « Les pâturages à Festuca eskia dans les Pyrénées ariégeoises, écologie. Composition floristique. Intérêt économique », Pirineos, 79-80 (Actes du cinquième congrès international d'études pyrénéennes, Jaca) : 159-170
Résumé CBN : l'article décrit la physionomie, la composition, la répartition et l'écologie des pelouses ouvertes à gispet des fortes pentes de soulane ("pelouses steppiques"). Il précise l'intérêt pastoral de ces formations pour les Pyrénées ariégeoises. Il mentionne d'anciennes pratiques de fauche (19e siècle)
CORRIOL Gilles, 2008, « Essai de clé typologique des groupements végétaux de Midi-Pyrénées et des Pyrénées françaises, I : introduction et pelouses acidophiles (Nardetea et Caricetea curvulae) », Monde des plantes, 495 : 3-13
Résumé auteur : Dans la perspective d'une typologie des habitats naturels de Midi-Pyrénées et des Pyrénées françaises, des clés de travail thématiques basées sur la littérature phytosociologique sont produites. Le présent article débute la série avec les classes des Nardetea strictae Rivas Goday et des Caricetea curvulae Br.-Bl.
GAUSSEN Henri, 1971, « Les gradins de Gispet aux Pyrénées », Bulletin de la Société Botanique de France, 118 (2) : 65-70, http://dx.doi.org/10.1080/00378941.1971.10838985
Résumé CBN : l'article analyse et interprète la physionomie en gradins des pelouses ouvertes. Selon l'auteur, cette disposition est due essentiellement au cheminement des troupeaux ovins.
GRUBER Michel,1975, " Les associations du Nardion Br.-Bl. 1926 en Pyrénées ariégeoises et catalanes ", Bulletin de la Société botanique de France 122 : 401-416
Résumé auteur : L'auteur étudie les pelouses s'intégrant à l'alliance du Nardion Br.-Bl. 1926 des Pyrénées ariégeoises et catalanes. Le Nard domine dans deux associations : le Selino-Nardetum Br.-Bl. 1948 mésohygrophyle, autour des lacs de montagne, et l'Alchemillo-Nardetum ass. nova mésophile, des replats et des fonds de vallées humides. La pelouse fermée à Festuca eskia Ram., localisée sur les versants exposés au nord, forme l'Androsaci-Festucetum eskiae ass. nova dont l'écologie est voisine des Nardaies et s'éloigne notablement du Campanulo-Festucetum eskiae Br.-Bl. 1948 des pentes eposées au midi. Le Trifolio-Alopecuretum gerardii Br.-Bl. 1948 est plutôt rare dans la dition et ses caractères écologiques sont assez semblables à ceux des combes à neige acidophiles.
MALAVAL-CASSAN Sandra, CORRIOL Gilles, LARGIER Gérard, 2005,"Festuca eskia Ramond ex DC. en Pyrénées françaises, bilan nomenclatural, taxonomique et écologique", Bull. Soc. Hist. Nat., Toulouse, 141-2 : 49-56
Résumé auteur : Espèce emblématique des montagnes pyrénéennes, le gispet (Festuca eskia Ramond ex DC.) est une graminée endémique des Pyrénées et des Monts Cantabriques, occupant de très larges surfaces dans les pentes raides et les éboulis. Publiée en 1805 par De Candolle (in LAMARCK et DE CANDOLLE), l’espèce est imputée à Ramond de Carbonnières, qui l’a récoltée en herbier dès 1793. Nous tenterons de faire le point sur la chorologie de l’espèce à la lumière d’informations en provenance des Carpathes roumaines où l’espèce aurait été vue par Klás√tersky√en 1922 (KRAJINA, 1933).
L’ensemble des taxons appartenant à la section Eskia Willk. dont Festuca eskia est le type ont fait l’objet de récentes études macro, micromorphologiques, voire moléculaires, permettant de préciser l’organisation de cette section. Les études morphologiques remettent en cause la distinction de deux variétés (var. orientalis et var. eskia) sur la chaîne pyrénéenne proposée par NÈGRE (1975). Elles permettent également de présenter les hybridations de Festuca eskia avec deux autres espèces de sa section : F. gautieri et F. quadriflora. Enfin, nous nous attacherons à présenter succinctement sa synécologie au long de la chaîne.
NEGRE R., 1974, "Nouvelle contribution à l'étude des gispetières pyrénéennes", Bol. Soc. Broteriana 48 : 209-251
NEGRE R., 1977, "Vue d'ensemble sur les pelouses à Festuca eskia et Festuca paniculata en Pyrénées", in Documents phytosociologiques vol. 1 : 189-194
NEGRE R., GHIGLIONE C., MARC P., 1986, "Productivité et valeur fourragère des gispetières pyrénéennes", Bulletin de la Société Botanique de France Vol. 133, Suppl. 1. Actes du Colloque International de Botanique Pyrénéenne (La Cabanasse) 3-5 juillet 1986 : 379-398
Résumé auteur : Les gispetières pyrénéennes se rattachent à cinq grandes associations dépendant directement de leur situation géographique dans la chaîne. Leur productivité et leur valeur fourragère sont déterminées par les caractères géomorphologiques des stations en relation avec le méso- et le micro-climat, tout autant que par la nature des espèces dominantes, dont la composition chimique varie avec les mêmes facteurs.
NEGRE R., SERVE L., 1979, "Projection dans les groupements à Festuca eskia en Pyrénées orientales", in Documents phytosociologiques vol. 4 : 731-756
PASCHE F., ARMAND M., GOUAUX P., LAMAZE T., PORNON A., 2004, “Are meadows with high ecological and patrimonial value endangered by heathland invasion in the French central Pyrenees?”, Biological Conservation 118 (2004) : 101–108, http://www.sciencedirect.com/science/article/pii/S0006320703003082
Résumé auteur : Due to their high ecological and patrimonial value Festuca eskia and Nardus stricta meadows andRhododendron ferrugineum heathlands need to be locally preserved in the Pyrenees. This could pose dilemmas to conservation managers if heathlands invade and replace the meadows. However, studies showing that the heathlands do actually invade meadows are lacking in the Pyrenees and the invasion hypothesis therefore remains to be proved. So, vegetation pattern changes in a particularly vulnerable site from the central Pyrenees was investigated by two means: (i) interpretation of stereo pairs of aerial photographs and (ii) analysis of the demography of Rhododendron populations in ten 150 m2 experimental plots. The main results from photo interpretation indicated that, during the period between 1954 and 1995, 15% of a 17 ha meadow area shifted to heathland. This corresponded with a 592 m2 year−1 colonization rate and to an overall front progression of heathland over meadows of 3–60 m. Demographic data showed that meadow colonization by heathland passed through successive phases but appeared to be a rather slow process that can take as long as 120–320 years. During the process the cover rate over the 150 m2 plots varied between 1 and 0.4 m2 year−1. Heathland extension does not seem to have been favored by the decrease in grazing pressure. Despite this, the progression of heathlands over meadows seems endangering the meadows. Finally we propose practices locally to limit the progression of Rhododendron.
Note de lecture (R. PRUNIER 2013) : L'étage de végétation subalpin des Pyrénées est constitué d'une mosaïque de pelouses, landes et forêts. Ce paysage est modelé par le climat et l’activité humaine depuis plus de 4000 ans. Le pâturage et les pratiques associées sont restés intenses jusqu'à la fin des années 1960, puis l'exode rural a modifiée l'équilibre entre les composantes du paysage. Dans la zone cœur du Parc national des Pyrénées, Pasche et al. (2003) ont étudié la dynamique des landes à Rhododendron ferrugineum, en contact avec des pelouses à Nard (Nardus stricta) ou à Gispet (Festuca eskia). Leur objectif a été de savoir si les pelouses sont en train de se convertir en lande, et de connaître la vitesse d'invasion. Cette étude met en évidence que la vitesse de colonisation du Rhododendron est assez lente et varie selon les cas, sur les 17 ha de pelouse étudiés 15 % ont été colonisés en 40 ans. L'analyse de la dynamique des populations de Rhododendron a permis de mettre en évidence quatre phases distinctes dans la colonisation, concernant l'arrivée de graines fondatrices, le recrutement de plantules, la multiplication végétative des individus, la fermeture du milieu. Ce processus de colonisation lent peut s'étaler sur une durée de 120 à 320 ans. L'extension observée s’effectue surtout à travers la multiplication végétative, la croissance des individus et la convergence des patches. En effet les plantules de rhododendron sont sensibles à la compétition face à la présence arbustes mais aussi face à un couvert herbacé dense. Dans cette étude, les parcelles faiblement pâturées présentent un recouvrement dense de Nard ou de Gispet et montrent la dynamique la plus lente. Ainsi, un pâturage peu intense permet de ralentir localement l'invasion du Rhododendron (Pasche et al., 2003). En définitive la réduction de la pression pastorale depuis 40 ans ne semble pas avoir favorisé l'extension de la lande sur le site étudié (Pasche et al., 2003). Selon leur nature, les activités pastorales ne participent pas toujours au maintien des milieux ouverts. Bien que le pâturage intense diminue survie des plantules ligneuses par abroutissement et déracinement, les effets négatifs sont largement compensés par la création de microsites favorables à l’installation du Rhododendron. Ainsi en contexte subalpin, le pâturage du bétail est susceptible de participer à convertir une pelouse en lande.
TOSCA Claude, LABROUE Louis, 1982, « Productivité primaire des pelouses subalpines et alpines : photosynthèse nette des espèces du fond prairial », in "Lazare J.J., Marty M., Dajor R. (Editeurs), 1984, Ecologie des milieux montagnards et de haute altitude" : 257-264
Résumé auteur : Au cours de l’été 1980 a été étudiée la photosynthèse nette des espèces qui constituent le fond prairial des pelouses supraforestières des Pyrénées centrales : Nardus stricta, Festuca rubra, Festuca eskia et Trifolium alpinum dans l’étage subalpin, Festuca scoparia, Elyna spicata, Carex curvula dans l’étage alpin. L’absorption de CO2 a été mesurée in situ à laie d’un analyseur à infrarouge en montage différentiel et à deux périodes de la saison estivale (juillet et septembre). A l’étage subalpin, les espèces présentent des capacités photosynthétiques très différentes si l’on rapporte l’activité au végétal, la moins active étant Festuca eskia, la plus active Festuca rubra, mais les rendements au m² de pelouse de ces espèces sont inversés du fait du pouvoir de recouvrement de chacune d’entre elles. A l’étage alpin, les espèces étudiées ont toutes des activités photosynthétiques très élevées, mais en raison du recouvrement très faible de ces espèces, les accroissements de biomasse sont du même ordre qu’à l’étage inférieur (entre 8 et 20 g de matière sèche par m² de pelouse et par jour). Quelle que soit l’altitude, on observe en septembre une activité nettement moindre qu’en juillet pour toutes les espèces, à l’exception de Festuca eskia. De même, ces espèces (sauf Nardus stricta et Festuca eskia) présentent une dépression de la photosynthèse sous l’action des forts éclairements de la mi-journée). Le fonctionnement observé des espèces d’origine méditerranéennes ne nous permet pas, pour l’instant, de les rattacher à des plantes de type C.
TOSCA Claude, 1975, « Les pelouses subalpines des Pyrénées centrales : leur valeur pastorale », Fourrages, 61 : 48-73, http://www.afpf-asso.fr/index/action/page/id/33/title/Les-articles/artic...,
Résumé auteur : Dans cet article, l'auteur rapporte les premiers résultats d'une étude de quelques pelouses subalpines des Pyrénées centrales. Quatre types physionomiques se partagent la majeure partie du territoire qui s'étage de 1.800 à 2.000 m d'altitude ; la détermination de leur valeur pastorale a été effectuée à partir des contributions spécifiques de chaque espèce végétale, en utilisant les indices de qualité de Delpech. Il apparaît ainsi que la connaissance des contributions spécifiques du fond prairial (fétuque rouge et trèfle alpin) permet de différencier dans la nardaie trois groupes de pâturages en relation avec les conditions écologiques stationnelles (exposition, pente, drainage). Cependant, et contrairement à ce qu'on observe avec la fétuque rouge, il n'existe pas de relation linéaire entre la valeur pastorale des herbages et la contribution spécifique du trèfle alpin ; en effet, la présence de cette espèce est conditionnée par le pH et l'humidité du sol. Par contre, la fétuque rouge participe à la colonisation de tous les sols, indépendamment des autres conditions écologiques, et elle entre en concurrence avec le nard. La connaissance des indices d'intensité d'exploitation permet de suggérer des possibilités d'amélioration de ces herbages par un aménagement du mode de pâture des troupeaux, en fonction du type de pelouse.
Données cartographiques
Le tableau et la carte ci-dessous montrent la répartition des surfaces cartographiées en « Pelouses pyrénéennes siliceuses à Festuca eskia » (UE 6140) dans le réseau régional de sites Natura 2000. Les surfaces portées dans la deuxième colone comprennent les polygones purs (un seul habitat) et les polygones mixtes (plusieurs habitats en mélange ou en mosaïque).
| Code Site Natura | Nom Site Natura | Surface (en ha) |
| FR7300827 | Vallée de l'Aston | 1 341,09 |
| FR7200744 | Massif de Sesques et de l'Ossau | 1 093,79 |
| FR7300880 FR7300881 |
Haute vallée d'Oô Haute vallée de la Pique |
1 026,23 |
| FR7300934 | Rioumajou et Moudang | 932,70 |
| FR7300926 | Ossoue, Aspé, Cestrède | 816,60 |
| FR7300935 | Haut-Louron : Aygues Tortes, Caillauas, Gourgs Blancs, Gorges de Clarabide, pics des Pichadères et d'Estiouère, montagne de Tramadits | 567,07 |
| FR7300931 | Lac Bleu Léviste | 534,13 |
| FR7300927 | Estaubé, Gavarnie, Troumouse et Barroude | 506,18 |
| FR7300932 | Liset de Hount Blanque | 477,51 |
| FR7200746 | Massif de l'Anie et d'Espelunguère | 385,61 |
| FR7300821 | Vallée de l'Isard, mail de Bulard, pics de Maubermé, de Serre-Haute et du Crabère | 375,05 |
| FR7300831 | Quérigut, Laurenti, Rabassolles, Balbonne, la Bruyante, haute vallée de l'Oriège | 324,23 |
| FR7300923 | Moun Né de Cauterets, pic de Cabaliros | 318,95 |
| FR7300822 | Vallée du Riberot et massif du Mont Valier | 286,67 |
| FR7200743 | Massif du Ger et du Lurien | 259,59 |
| FR7300925 | Gaube, Vignemale | 205,71 |
| FR7300883 | Haute vallée de la Garonne | 172,74 |
| FR7300924 | Péguère, Barbat, Cambalès | 130,83 |
| FR7300921 | Gabizos (et vallée d'Arrens, versant sud-est du Gabizos) | 121,42 |
| FR7300930 | Barèges, Ayré, Piquette | 95,06 |
| FR7300929 | Néouvielle | 60,01 |
| FR7200745 | Massif du Montagnon | 16,59 |
| FR7200750 | Montagnes de la Haute Soule | 6,81 |
| FR7200742 | Massif du Moulle de Jaout | 5,23 |
| FR9101470 | Haute vallée de l'Aude et bassin de l'Aiguette | 0,55 |
| Total 6140 | 10 060,36 |
Les pelouses denses à Festuca eskia se rencontrent exclusivement dans les sites de montagne de la chaîne pyrénéenne.
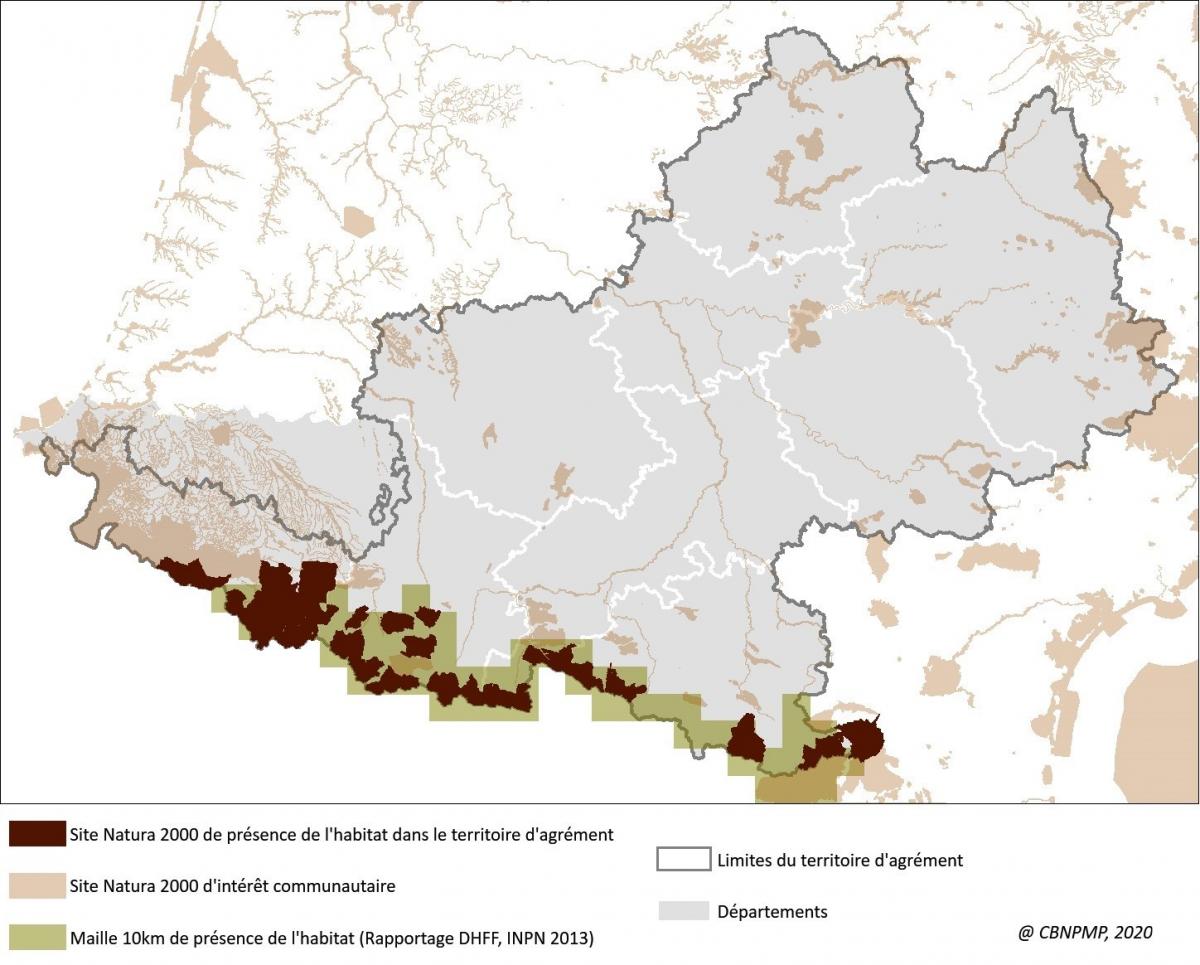
 La taille moyenne des unités cartographiées en pelouse dense à Festuca eskia est de 6 ha, les gispetières pures représentant 1/4 des surfaces.
La taille moyenne des unités cartographiées en pelouse dense à Festuca eskia est de 6 ha, les gispetières pures représentant 1/4 des surfaces.
Les habitats en mélange sont principalement des landes alpines et montagnardes, des nardaies et des éboulis ou pentes rocheuses sur substrat acide. Le mélange avec les landes révèle, pour partie, une relation dynamique en lien avec une évolution des conditions d'exploitation pastorale.
État de conservation
Données issues du rapportage SPN-2015 // MNHN et Fédération des CBN
Selon la Directive Habitats, un état de conservation est jugé favorable « lorsque l’aire de répartition naturelle ainsi que les superficies couvertes par l’habitat au sein de cette aire sont stables ou en extension, et la structure et les fonctions spécifiques nécessaires à son maintien à long terme existent et sont susceptibles de perdurer dans un avenir prévisible, et l’état de conservation des espèces qui lui sont typiques est favorable. ». Tous les 6 ans, un bilan est réalisé à l’échelle nationale et européenne sur la base de 4 indicateurs : aire de répartition, surfaces, structure et fonctions.


Groupes de gestion identifiés
| Groupe Fonctionnel A | Syntaxon | CORINE BIOTOPE | EUR 28 | CH |
|---|---|---|---|---|
 | Ranunculo pyrenaei-Festucetum eskiae Nègre 1969 | 36.314 | 6140 | 6140-1 |
![[6140-1] Gispetière dense (vallée d'Ossoue 65 - photo R. Prunier)](https://natura2000.cbnpmp.fr/sites/default/files/gispetiere_denserobin_ossoue_27juin13_dscn1383.jpg)
![[6140-2] Les gispetières fermées se développent sur des sols relativement profonds, là où la couverture neigeuse peut constituer un abri vis à vis du vent et des fortes gelées. Ici, seules quelques petites portions de versant (sous le sentier, bas de pente...) remplissent les conditions propices à l'installation de l'habitat (Oulettes de Gaube 65 - photo C. Brau-Nogué)](https://natura2000.cbnpmp.fr/sites/default/files/gispetiere_versant_vignemale_6aout15_p1010417.jpg)
![[6140-3] Gispetière semi-fermée. Dans la forme typique de gispetière fermée d'ombrée, le recouvrement de la végétation dépasse souvent 90%. Néanmoins, la part de sol nu peut aller jusqu'à 25 voire 30% : l'habitat prend alors une physionomie de pelouse semi-ouverte qui se rapproche de celle des gispetières en gradins de versant sud (code corine 36.332 - hors DH). Les formes intermédiaires (mosaïques ou transitions) sont fréquentes mais ces deux types de gispetières n'ont a priori pas de lien dynamique.](https://natura2000.cbnpmp.fr/sites/default/files/gispetiere_semi-fermee_robin_ossoue_27juin13_dscn1408.jpg)
![[6140-4] Pelouse à Gispet mêlée de Callune et de Myrtille. Cette mosaïque semble aujourd'hui en équilibre mais pourrait évoluer vers une lande si la durée d'enneigement ou la pression de pâturage diminuent durablement (col de Pailhères 11 - photo R. Prunier)](https://natura2000.cbnpmp.fr/sites/default/files/gispet_myrtille_callune_ascou_27juin13dscf2260.jpg)
![[6140-5] Un enneigement prolongé est nécessaire au maintien des gispetières denses. Ici, l'habitat se retrouve au niveau d'une congère ; en dehors de cette zone particulière c'est une lande à callune et myrtille qui domine (col de Pailhères 11 - photo R. Prunier)](https://natura2000.cbnpmp.fr/sites/default/files/gispet_sous_congere_robin_ascou_27juin13dscn1108.jpg)
![[6140-6] Touffe de Gispet pâturée. Le prélèvement se limite généralement à l'extrémité des limbes (vallon d'Arizes 65 - photo C. Brau-Nogué)](https://natura2000.cbnpmp.fr/sites/default/files/gispet_touffe_paturee_arizes_12aout15p1010621.jpg)
![[6140-7] Mosaïque pelouse-lande. Les pelouses fermées à Gispet sont souvent associées à des habitats de lande, en particulier les landes à Rhododendron et Myrtille. Cette proximité peut venir d'une hétérogénéité des conditions stationnelles : on remarque ici que les ligneux occupent préférentiellement les épaulements et les parties plus rocailleuses. Mais elle peut aussi révéler une situation dynamique avec une progression plus ou moins rapide de la lande sur l'habitat de pelouse. La distinction entre les deux phénomènes n'est pas toujours facile à établir. (vallée d'Ossoue 65 - photo C. Brau-Nogué)](https://natura2000.cbnpmp.fr/sites/default/files/gispetiere-rhodoraie_ossoue_22juil13p1080083.jpg)
![[6140-8] Malgré le pâturage bovin, la dynamique de fermeture semble ici bien engagée. Les premiers arbres (Pins à crochets, Bouleaux, ...) sont déjà en place : à moyen terme, la lande à Rhododendron sera elle-même très certainement remplacée par une forêt de montagne. (Vallée d'Ossoue 65 - photo C. Brau-Nogué))](https://natura2000.cbnpmp.fr/sites/default/files/gispetiere-rhodoraie_vaches_ossoue_22juil13p1080086.jpg)